Ana was 22 years old when she found out she had it… Hepatitis C! Unlike others her age, she hadn’t yet had a boyfriend, gotten tattoos in non sterile conditions, damaged her liver by excessive drinking in college, or shared needles to inject drugs, all of which are ways one can get the disease (1). All she had done was receive a kidney (a solid organ) from her uncle who was born in 1962 in Japan, one of the few industrialized countries with high Hepatitis C rates (2). She first noticed the massive bruises on her arms and legs 6 months ago followed by the yellowing of her eyes (jaundice). Her doctor thought it was her body rejecting the kidney and causing her to have reduced platelets but after getting a biopsy, it turned out her liver had become cirrhotic and was failing. Cirrhosis is a chronic disease that leads to inflammation which, over time, replaces healthy liver cells with scar tissue. It takes about 20 to 30 years for this to happen (or faster if alcohol is drunk or if someone has HIV). The Hepatitis C virus had entered into her body through her uncle’s kidney transplant which had destroyed her liver. She had lupus which destroyed her original kidneys and resulted in the transplant from her dear uncle.

Figure 1. Disease spectrum of alcoholic liver disease. Image from Singal et al. 2017.
Hepatitis C is a viral infection that causes liver inflammation, sometimes leading to serious liver damage including cirrhosis and liver cancer (see Figure 1). The hepatitis C virus (HCV) spreads through contaminated blood during high-risk behaviours such as injection drug use, but can also spread through solid organ donation. After the incubation stage (14 to 80 days), most people clear the virus within the first 6 months of an acute infection. If this doesn’t happen, the chronic infection causes constant inflammation that, over time, replaces healthy liver cells with scar tissue and increases the risk of liver cancer.
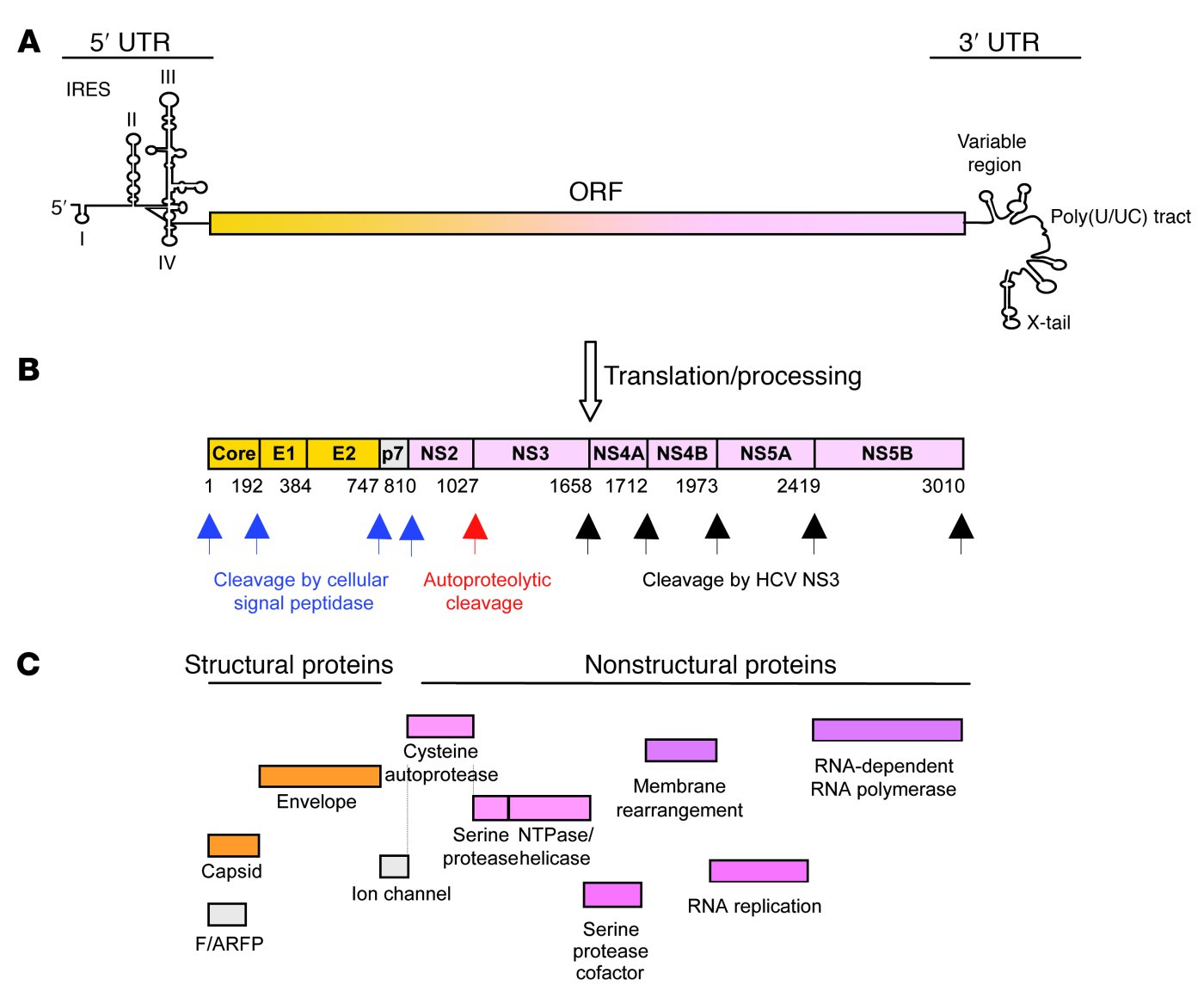
Figure 2. (a) The single-stranded RNA genome encodes a long open reading frame (ORF) flanked by 2 UTRs, which contain signals for viral protein and RNA synthesis and the coordination of both processes. Translation is initiated through an internal ribosome entry site (IRES) in the 5’ UTR. U, uridine; C, cytidine. (b) The translated polyprotein is cotranslationally and posttranslationally processed by cellular and viral proteases. Numbers below the polyprotein indicate the amino acid positions of the cleavage sites. (c) Function of the resulting 10 structural and nonstructural proteins. A frameshift (F) protein is translated from a short alternate reading frame (ARF). Image from Rehermann B., 2009.
HCV is a small (55-65 nm in size) single stranded RNA virus. It has a single-stranded RNA genome which consists of a single open reading frame (ORF) sandwiched between the untranslated 5’ and 3’ ends of the RNA (ie. 5’UTR and 3’UTR). The 5’ UTR has a ribosomal binding site or internal ribosome entry site (IRES) (see Figure 2a). This IRES initiates the translation in a cap-independent manner of a very long protein containing about 3,000 amino acids. In regular RNA processing, pre-RNA is transcribed from DNA. After transcription, a 5’cap as well as a poly(A) tail are both added to the mRNA to protect it from degradation during the RNA processing. When the RNA is mature (mRNA), it then gets translated into a polypeptide (see Figure 3). The HCV, however, is initiated by a different mechanism than this 5’cap-binding model. It relies on the direct binding of the 40S ribosomal subunit by the IRES.

Figure 3. RNA processing to be translated into a peptide. Image from in Slideshare, 2007.
HCV was able to infect Ana’s cells using a Type 3 IRES (see Figure 4). IRESs are important RNA-based translation initiation signals, critical for infection by many pathogenic viruses. IRES RNA functionally replaces several large initiation factor proteins by directly recruiting the 43S particle: the 40S ribosomal subunit and eukaryotic initiation factor 3 (eIF3) (see Figure 4).

Figure 4. HCV core IRES Type 3. Image from Svitkin et al. 2015
Three independently folded structural domains in the IRES RNA make intimate contacts to two purified components of the 43S particle. Precise functioning of the HCV IRES requires the specific interaction of its domains with ribosomal subunits and a subset of eukaryotic translation initiation factors (eIFs). eIF1A is necessary for efficient activity where it stabilizes tRNA binding and inspects the codon-anticodon interaction.
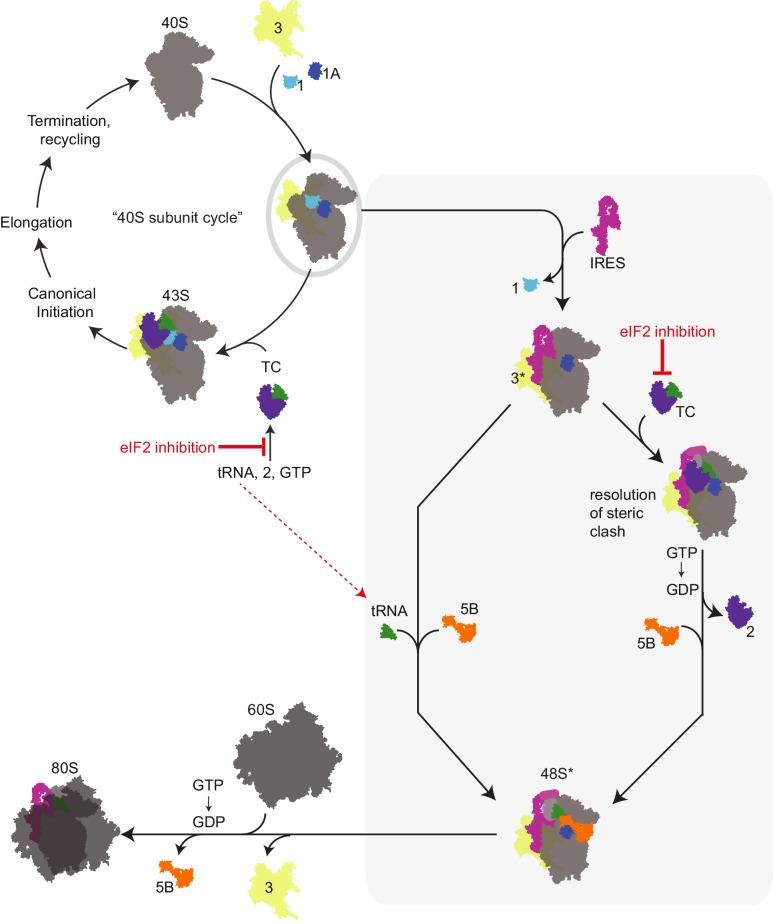
Figure 5. HCV IRES and type 3 IRESs recruits a pre-assembled “ternary complex-deficient” (‘TC-deficient’) preinitiation complex (PIC) containing eIF1, 1A, and 3 (grey oval), which is an intermediate in the normal 40S recycling process. IRES binding to this PIC partially remodels it to include displacing eIF3 (yellow, ‘3*’) and either passively or actively releasing eIF1. This remodeled PIC is used under both eIF2 active and inactive conditions (grey shaded box). When active eIF2 is abundant, delivery of tRNA through the TC presumably requires movement of IRES domain II to an alternate position to resolve a steric clash. If eIF2 is inhibited, this would have the effect of increasing the amount of available ‘TC-deficient’ PICs and perhaps free tRNA. The eIF3 is lost and the large subunit, 60S binds to the small subunit to create 80S. 80S is then able to go ahead and translate the RNA. Image from Jafaar et al., 2016.
HCV IRES binds 40S in such a manner that AUG initiator codon is positioned in the ribosomal P-site (peptidyl site, the second binding site for tRNA in the ribosome). As a result, scanning factors eIF1 and eIF1A are can be used for the HCV translation, as well as other components which are generally required for mRNA binding and unwinding of 5'UTR. Initiator tRNA is delivered either by eIF2 or, in stress conditions when eIF2 is inactivated, by eIF2A, eIF2D, or possibly eIF5B, a homologue of prokaryotic IF2 protein. Thus allowing the RNA to be translated into protein.
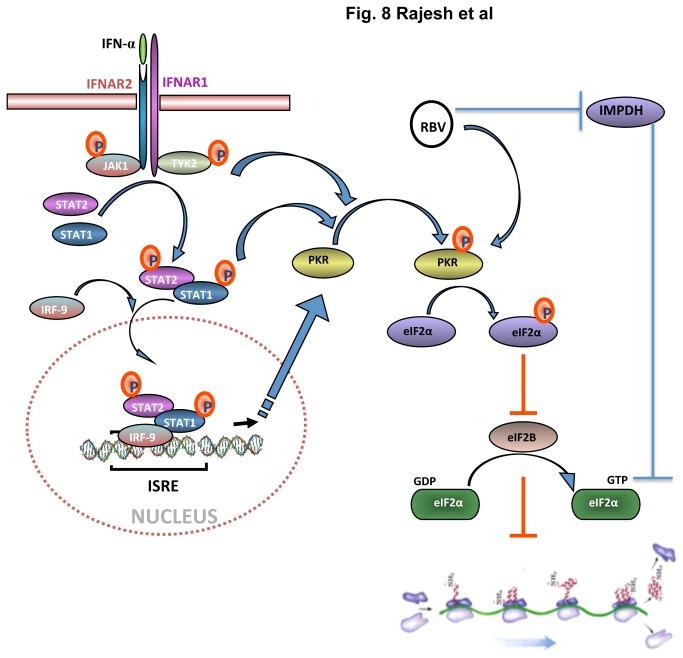
Ana didn’t choose to have lupus and didn’t choose to have Hep C now. But it happens. Luckily, she’s heard there’s a new drug combination (Interferon & Ribavirin) that can cure her HCV and even reverse her liver failure even though it does not directly target IRESs (see Figure 6).
About the Author

Chukwuamaka Onyewadume is a Senior at Mount Holyoke College majoring in Biochemistry. She discovered her love for research working in Professor Katie Berry’s lab at Mount Holyoke College where she is studying ProQ-RNA interactions in E.coli. From Nigeria, Chukwuamaka plans on pursuing an MD-PhD and specialising as a Pediatric Intensivist.
References
(1) Hepatitis questions and answers for the community. Centers for Disease Control and Prevention. https://www.cdc.gov/hepatitis/hcv/cfaq.htm#D5
(2) Petruzziello, A., Marigliano, S., Loquercio, G., Cozzolino, A., and Cacciapuoti, C.Petruzziello, Arnolfo et al. “Global epidemiology of hepatitis C virus infection: An up-date of the distribution and circulation of hepatitis C virus genotypes.” World journal of gastroenterology vol. 22,34 (2016): 7824-40. doi:10.3748/wjg.v22.i34.7824 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5016383/
(3) Rehermann B. 2019. Hepatitis C virus versus innate and adaptive immune responses: a tale of coevolution and coexistence. The Journal of Clinical Investigation. 2009;119(7):1745-1754. https://doi.org/10.1172/JCI39133
(4) Jubin R. 2001. "Hepatitis C IRES: translating translation into a therapeutic target". Curr. Opin. Mol. Ther. 3(3):278–87. PMID 11497352.
(5) Central Dogma and Protein synthesis. In Slideshare, 2007. https://www.slideshare.net/cgales/central-dogma-and-protein-synthesis
(6) Svitkin, Yuri & Siddiqui, Nadeem & Sonenberg, Nahum. (2015). Protein Synthesis Initiation in Eukaryotes: IRES-mediated Internal Initiation.
(7) Jaafar, Z.A., Oguro, A., Nakamura, Y., Kieft, J.S. 2016. “Translation initiation by the hepatitis C virus IRES requires eIF1A and ribosomal complex remodeling.” eLife vol. 5 e21198. 23 Dec. 2016, doi:10.7554/eLife.21198
(8) Panigrahi, R., Hazari, S., Chandra, S., Chandra, P.K., Datta, S., Kurt, R., Cameron, C.E., Huang, Z., Zhang, H., Garry, R.F., Balart, L.A., Dash, S. 2013. “Interferon and ribavirin combination treatment synergistically inhibit HCV internal ribosome entry site mediated translation at the level of polyribosome formation.” PloS one vol. 8,8 e72791. 23 Aug. 2013, doi:10.1371/journal.pone.0072791
Comments
Post a Comment