Virus-encoded Circular RNAs:
A brand-new class of gene regulators
What is a normal shape of general RNAs you have learned about before? Are all types of RNAs linear as we often think? The answer is NO!
Different from other members in the RNA-family, circular RNA (circRNA) is a novel class of RNA that is not linear. Instead, it forms a covalently closed continuous loop in which the 3'end and 5'end in an RNA molecule are joined together. In eukaryotes, pre-mRNA goes through processing which the 5'end is capped with 7MetG and the 3'end is added with a lot of adenines. With circRNAs, the absence of the two ends as well as the polyadenylated tail makes circRNAs resistant to degradation and more stable than most linear RNAs in cells. With this distinct shape, circRNAs display many original properties that other RNAs don't have. Therefore, circRNA is one of the most actively fields in RNA-targeted therapeutic treatments of research in recent years.
CircRNA is more represented in eukaryotic RNA molecules than in prokaryotic ones, and varies widely in different species. In previous years, scientists thought that circRNAs are just by-products of splicing errors with few functions. However, more and more studies have been showing that circRNAs do have potentially important roles in gene regulation, including the titration of microRNAs (miRNAs), regulation of transcription and interference with pre-mRNA splicing. Therefore, circular RNAs were officially declared as a new class of RNAs in 2016.
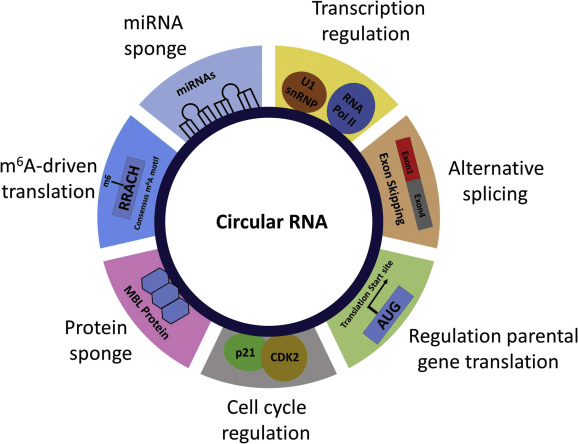
Figure 1. Multiple functions of circRNAs in molecular biology.
Figure retrieved from: Huang et.al, 2017
Having a different structure than better known RNAs, circRNAs open a question of interest: What are potential mechanisms that can lead to circular RNA formation?
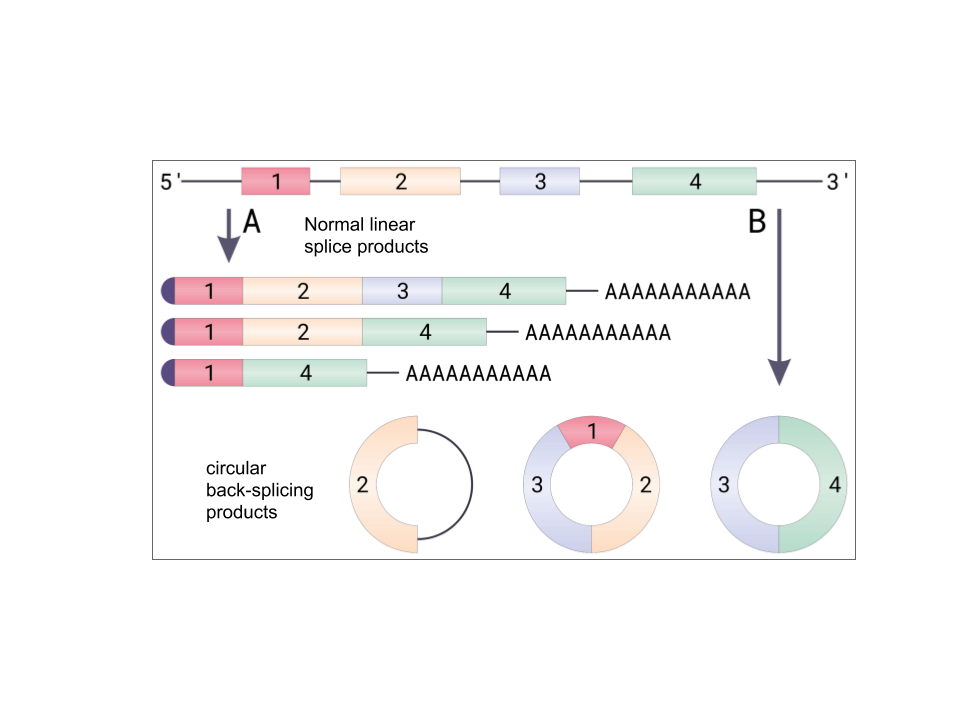
Figure 2. Circular RNA formation from pre-mRNA via backsplicing event. (A) Canonical splicing of mRNA with different splice variants. All resulting mature mRNAs have 5'end cap and 3'end poly-A tail. (B) Circular formation can have one or more exons, sometimes introns are still retaining in the final circRNA. Figure retrieved from Wikipedia.
In molecular biology, pre-mRNA made after transcription needs to go through RNA processing in order to form a mature mRNA. One of the mRNA processing steps is called splicing, which introns are removed and exons are joined together in a linear way. However, using a different spliceosomal machinery, circRNA is produced by back-splicing circulization. As researchers are still uncovering how the spliceosome is involved in back-splicing, Chen et.al (2016) proposed a mechanism that spliceosomal small nuclear ribonucleoproteins (snRNPs) can be assembled on the precursor mRNA (pre-mRNA). The 5'end of mRNA acts a donor site of an exon and is joined to the 3'end (acceptor site) to form a circular RNA (Figure 3b). With this multiple splicing sites, competition for different exons joining leads to the formation of different circRNAs from just a single gene (Figure 2).

Figure 3. Proposed mechanism of circRNA formation.
Figure retrieved from Chen et.al (2016)
CircRNAs are often expressed at low levels in eukaryotic transcripts. Therefore, for many years they have been overlooked by the assumption that a majority of circRNAs are just inactive splicing by-products. However, scientists discovered around 50 genes that have circRNAs expressed more abundantly than their linear isoforms in specific cell lines. Recently, dozens of circRNAs have been experimentally elucidated to be highly expressed in a cell type, tissue and even developmental-stage-specific manner.
More than 10,000 different circular RNAs have been identified in fruit flies, worms, mice and humans. They are more recently to be found in plants, fungi and protists. However, whether or not viruses can encode circRNAs remained unknown for many years. The Epstein-Barr virus (EBV) attracted a group of researchers at Sun Yat-sen University (Guangzhou, China) because EBV is widely known to produce non-coding RNAs such as small RNAs, miRNAs and long non-coding RNAs. They looked deeply into different EBV transcripts that both contain introns and exons to find out the potential transcripts where splicing can occur to generate circular RNAs.

Figure 4. Epstein-Barr virus (EBV) is a large genomic DNA virus of the herpesvirus family that infects over 90% of adults worldwide. Figure retrieved from Rawls, 2017
Let me introduce you to the Epstein-Barr virus: its genome is approximately 170 kb, encoding more than 80 proteins. There were approximately 143,000 deaths worldwide in 2010 attributed to these EBV-associated malignancies, representing 1.8% of all cancer deaths. This makes EBV is a key player in chronic illnesses. Some threatening diseases include lymphoid and epithelial malignancies, nasopharyngeal carcinoma and even autoimmune disorders such as multiple sclerosis. Therefore, studying how circRNAs regulate gene expression in EBV could open a new era of developing RNA-targeted therapeutic treatments for many different diseases.
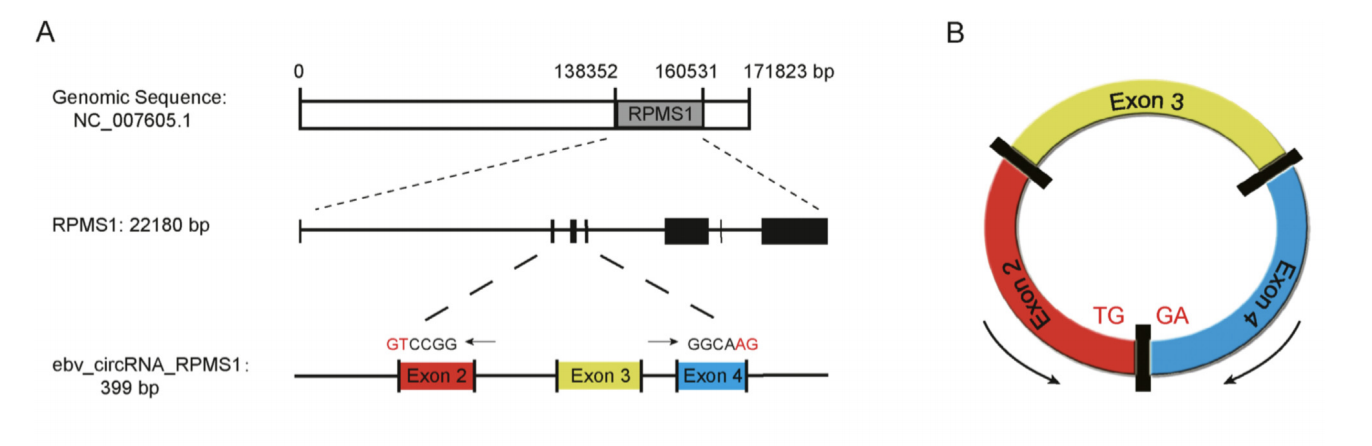
Figure 5. Schematic illustration of ebv-circRPMS1. (A) Genomic location of the RPMS1 gene and ebv-circRPMS1. (B) Schematic illustration showing the circularization of RPMS1 exons 2–4 forming ebv-circRPMS1. Figure retrieved from Huang et.al, 2019.
Huang et.al (2019) has found that the candidate circRNA has a head-to-tail splicing from the RPMS1 gene (Figure 5A). The circRNA is 399 base pairs out of 22180 base pairs in RPMS1 gene, denoted as ebv-circRNA-RPMS1. Ebv-circRPMS1 was localized both in cytoplasm and nuclei of the EBV-infected cells of human carcinoma tissues. It was found that the total RNA existing in circular form without a polyadenylated tail (ebv-circRPMS1) was resistant to digestion by RNase R exonuclease. Interestingly, this character has been well-established for circular RNAs in plants, mice and human experimental models in previous researches.
Ebv-circRPMS1 also target other non-coding RNAs, such microRNAs. MicroRNAs (miRNAs) are a class of small non-coding RNAs, which are about 20-22 nts in length and involved in gene regulation at a post-transcriptional level. Ebv-circRPMS1 can create continuous miRNA loss of function in RNA-guided gene regulation, known as miRNA sponge. It binds to both human and EBV miRNAs and sequester miRNAs from their targets, thus regulating the expression of other genes. Over 11 of 14 human miRNAs have been downregulated significantly after the overexpression of ebv-circRPMS1. Another class of circRNA, Cdr1, has been discovered to interact with miRNA-7 and miRNA-671 in human and mouse brains. Researchers found that when a mammalian circular RNA locus was lost, miRNA deregulation occurs that results in different brain impairment.

Figure 6. Many ongoing studies have been conducted to discover the association of circular RNAs with different diseases
Due to their covalently closed structure and high stability, circular RNAs have been confirmed to exhibit unique characteristics and functions in the RNA family. In fact, circRNAs have been an active field of research to build an RNA-targeted therapeutic treatments in many institutions around the world. The association of circRNA in many different pathways of gene regulation was confirmed in a wide range of human diseases such as gastric cancer, colon cancer, and neurodegenerative diseases. This is an exciting news to the future era of exploiting circRNAs for the development of useful biomarkers for diagnosis and prognosis, as well as the discovery of novel RNA-based therapies.
Written by Anh M. Tran’ 20
April 6th, 2019

About the Author: Anh M. Tran is a junior, majoring in Biochemistry at Mount Holyoke College. She comes from Ho Chi Minh city, Vietnam. Anh has a strong interest in studying the applications of RNAs in clinical studies, especial the gene regulatory functions of different RNAs in treating diseases. Hope you will enjoy this post about circular RNAs, and please feel free to reach me at tran28a@mtholyoke.edu if you have any questions!
References
Chen, L. L. (2016). The biogenesis and emerging roles of circular RNAs. Nature reviews Molecular cell biology, 17(4), 205.
Huang, G., Li, S., Yang, N., Zou, Y., Zheng, D., & Xiao, T. (2017). Recent progress in circular RNAs in human cancers. Cancer letters, 404, 8-18.
Huang, J. T., Chen, J. N., Gong, L. P., Bi, Y. H., Liang, J., Zhou, L., ... & Shao, C. K. (2019). Identification of virus-encoded circular RNA. Virology.
Jeck, W. R., Sorrentino, J. A., Wang, K., Slevin, M. K., Burd, C. E., Liu, J., ... & Sharpless, N. E. (2013). Circular RNAs are abundant, conserved, and associated with ALU repeats. Rna, 19(2), 141-157.
Li, X., Yang, L., & Chen, L. L. (2018). The biogenesis, functions, and challenges of circular RNAs. Molecular cell.
Rickinson, A., Kieff, E., 2007. Epstein-Barr virus. In: Knipe, D., Howley, P. (Eds.), Fields Virology, Fifth ed. Lippincott Williams & Wilkins, Philadelphia, pp. 2655–2700.
Salzman, J., Chen, R. E., Olsen, M. N., Wang, P. L., & Brown, P. O. (2013). Cell-type specific features of circular RNA expression. PLoS genetics, 9(9), e1003777.
Comments
Post a Comment