Summary of main points
- Retrotransposons are a type of mobile genetic element (MGE) that can copy and paste themselves multiple times throughout the genome using an RNA intermediate.
- Retrotransposons are widely considered to be harmful and bad for cells. They resemble retroviruses and our cells have developed mechanisms to protect against retrotransposition.
- The prevalence of retrotransposons throughout the tree of life suggests they are evolutionary important. This contrasts the negative effects that can cause in cells.
- Retrotransposons are suggested to generate more genetic diversity locally and globally in a genome, which is consistent with their prevalence in humans among other species.
- The glossary at the end of the post may have some helpful information should you need it :)
 |
| Figure 1: The mechanism of DNA transposition. |
In the past ten years alone, the study of mobile genetic elements (MGEs), or DNA sequences that can move around within a genome or between species’ genomes, has expanded exponentially. One class of mobile genetic elements is called transposons, which are MGEs with the potential to copy and paste themselves multiple times throughout a genome. The two major types of transposons, Class I and Class II each have a different mechanism by which they accomplish moving around in the human genome. Class II transposons or DNA transposons are pinched out from a sequence of DNA with the help of enzymes, and then inserted into a target site somewhere else in the genome (Figure 1).
DNA transposons are generally less studied than Class I transposons or retrotransposons (see Glossary at the end of post), which use a similar mechanism with one key difference: Class I transposons use an RNA intermediate to be copied and pasted, whereas Class II transposons lack such an intermediate. In other words, retrotransposons go from DNA → RNA → DNA, but DNA transposons go from DNA → DNA. The ratio of Class I to Class II transposons vary according to species, which hints that they may be involved with different cellular pathways (Figure 2).
 |
| Figure 2: The relative ratios of DNA transposons to retrotransposons in various species. This graph shows the contribution of DNA transposons and retrotransposons in percentage relative to the total number of transposable elements in each species. (Sc: Saccharomyces cerevisiae; Sp: Schizosaccharomyces pombe; Hs: Homo sapiens; Mm: Mus musculus; Os: Oryza sativa; Ce: Caenorhabditis elegans; Dm: Drosophila melanogaster; Ag: Anopheles gambiae, malaria mosquito; Aa: Aedes aegypti, yellow fever mosquito; Eh: Entamoeba histolytica; Ei: Entamoeba invadens; Tv: Trichomonas vaginalis.) |
A prominent subclass of retrotransposons that I will explore more in-depth throughout this post are Long Interspersed Elements (LINEs). LINE-1 (L1) represents the most abundant family of LINEs, but the majority of L1s are described as not-functional given they cannot be transcribed and/or translated. The structure of human L1 is shown in Figure 3.
 |
| Figure 3: The structure of human L1. |
The key component of the retrotransposon are its open reading frames (ORFs) of the retrotransposon. ORFs are simply the parts of the DNA that will eventually be translated by the ribosome once the DNA is converted to RNA. Human L1 has three ORFs, which are described in the following table (Figure 4).
The mechanism of L1 transposition is as follows (Figure 5):
Historically, retrotransposons have been associated with cellular dysfunction and the disruption of metabolic pathways that regulate cell life. They have been linked to disease phenotypes including tumour formation, cancer progression, blood clotting disorders, and Type 2 diabetes. Furthermore, an accumulation of L1 DNA in the cytoplasm initiates an immune response that stimulates inflammation. This L1-induced inflammation has been implicated in Alzheimer’s disease, Parkinson’s disease, arthritis, and many other pathologies. For these reasons, retrotransposons pose a risk to living cells, enabling evolution to equip our cells with some safety mechanisms against retrotransposition.
These mechanisms come in many flavours, but the most common target used to prevent retrotransposition is the RTase enzyme. You may have heard of RTase before, most likely
context of one of the most widespread viral pathogens, the human immunodeficiency virus (HIV). Similarly to retrotransposon activity, when HIV infects a cell, it injects a RTase enzyme into the cytoplasm in addition to its viral RNA genome. One of our cells’ protection mechanisms involves the use of a class of proteins called APOBEC (Figure 6), which physically inhibits the RTase enzyme, limiting retrotransposition by preventing L1 DNA from being made from L1 RNA.
Interestingly, APOBEC has also been shown to inhibit HIV’s RTase, though HIV deploys a special protein called viral infectivity factor (Vif) to counteract APOBEC’s inhibiting activity. HIV strains that are Vif-deficient are controllable using APOBEC proteins, which limits both HIV and L1 retrotransposition. However, most HIV strains contain Vif, blocking the activity of APOBEC and therefore enabling their own and L1 retrotransposition at the same time.
While it may seem like transposons have quite the notorious reputation, there is an evolutionary catch: transposons are present in genomes of virtually every organism, and in humans, transposons are conservatively estimated to comprise 45% of our DNA, 42% of which are retrotransposons. Why would transposons be so prevalent if they are so deleterious?
This remarkable prevalence of transposons in organisms and their tolerated accumulation in the human genome has led scientists to one reasonable hypothesis: retrotransposition greatly increases both local and global genetic diversity in a heritable way. When a retrotransposon is inserted into a target site somewhere else in the genome, there are some sticky end gaps that need to be filled in by a DNA polymerase enzyme (Figure 7).
This process often causes mutations that can have effects on downstream genes. Frequently, retrotransposition occurs in intronic DNA sequences that don’t code for any gene product such as RNA or a protein, but gene expression may still be affected. For example, if retrotransposition occurs in or near intronic sequences that regulate gene expression such as promoters or repressors, they have the potential to alter patterns of gene expression. Most significantly, these changes are heritable because they take place inside our DNA, and therefore can be acted upon by natural selection.
There is much about retrotransposition that we still do not understand, largely because retrotransposition is often limited to non-coding DNA. With only 1.2% of our DNA actually coding for RNA or proteins, the remaining 98.8% used to be referred to as “junk DNA” as it is considered non-coding. However, the rise of a new field of biology that studies so-called junk DNA has both challenged and aided in our understanding of gene expression and epigenetics. Retrotransposons are central to this area of knowledge, and their recent investigation has provided many platforms for developing new disease therapies. Perhaps they shouldn’t have such a bad reputation after all?
Glossary
 |
| Figure 4: The proteins encoded by each ORF of human L1. |
The mechanism of L1 transposition is as follows (Figure 5):
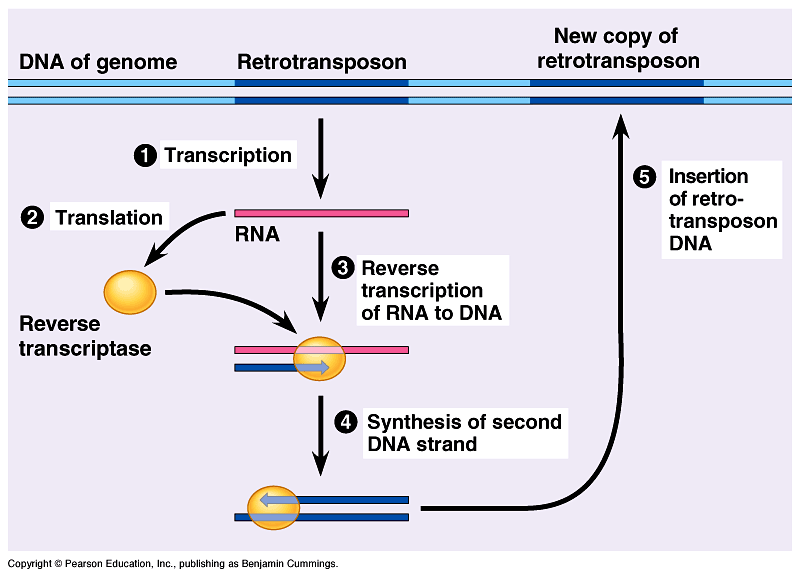 |
| Figure 5: The mechanism of L1 retrotransposition. |
- L1 DNA is transcribed into L1 messenger RNA (mRNA) by RNA Polymerase II.
- L1 mRNA is moved out of the nucleus into the cytoplasm and translated into the nuclear chaperone protein, exonuclease, and RTase enzymes.
- RTase begins to convert L1 mRNA in the cytoplasm back into two sequences of complementary L1 DNA.
- The nuclear chaperone protein complexes with the newly created L1 DNA and the exonuclease and re-enters the nucleus. Here, the exonuclease cuts a target site in the host-genome and the L1 DNA is inserted at the point where the host DNA was cleaved.
Historically, retrotransposons have been associated with cellular dysfunction and the disruption of metabolic pathways that regulate cell life. They have been linked to disease phenotypes including tumour formation, cancer progression, blood clotting disorders, and Type 2 diabetes. Furthermore, an accumulation of L1 DNA in the cytoplasm initiates an immune response that stimulates inflammation. This L1-induced inflammation has been implicated in Alzheimer’s disease, Parkinson’s disease, arthritis, and many other pathologies. For these reasons, retrotransposons pose a risk to living cells, enabling evolution to equip our cells with some safety mechanisms against retrotransposition.
These mechanisms come in many flavours, but the most common target used to prevent retrotransposition is the RTase enzyme. You may have heard of RTase before, most likely
 |
Figure 6: The general structure of an APOBEC protein.
|
Interestingly, APOBEC has also been shown to inhibit HIV’s RTase, though HIV deploys a special protein called viral infectivity factor (Vif) to counteract APOBEC’s inhibiting activity. HIV strains that are Vif-deficient are controllable using APOBEC proteins, which limits both HIV and L1 retrotransposition. However, most HIV strains contain Vif, blocking the activity of APOBEC and therefore enabling their own and L1 retrotransposition at the same time.
While it may seem like transposons have quite the notorious reputation, there is an evolutionary catch: transposons are present in genomes of virtually every organism, and in humans, transposons are conservatively estimated to comprise 45% of our DNA, 42% of which are retrotransposons. Why would transposons be so prevalent if they are so deleterious?
This remarkable prevalence of transposons in organisms and their tolerated accumulation in the human genome has led scientists to one reasonable hypothesis: retrotransposition greatly increases both local and global genetic diversity in a heritable way. When a retrotransposon is inserted into a target site somewhere else in the genome, there are some sticky end gaps that need to be filled in by a DNA polymerase enzyme (Figure 7).
 |
| Figure 7: A common mechanism by which retrotransposition can generate mutations. |
There is much about retrotransposition that we still do not understand, largely because retrotransposition is often limited to non-coding DNA. With only 1.2% of our DNA actually coding for RNA or proteins, the remaining 98.8% used to be referred to as “junk DNA” as it is considered non-coding. However, the rise of a new field of biology that studies so-called junk DNA has both challenged and aided in our understanding of gene expression and epigenetics. Retrotransposons are central to this area of knowledge, and their recent investigation has provided many platforms for developing new disease therapies. Perhaps they shouldn’t have such a bad reputation after all?
Glossary
- Mobile genetic elements (MGEs): genetic materials that can be transferred from one species to another or can be shuffled around or copied and pasted multiple times within one genome.
- Transposon: a segment of DNA that can be copied and pasted within one genome.
- Class I transposon: a retrotransposon that can copy and paste itself multiple times throughout the genome using an RNA intermediate. Goes from DNA → RNA → DNA.
- Class II transposon: a DNA transposon that can cut and paste itself multiple times throughout the genome. Goes from DNA → DNA.
- Open reading frame (ORF): the part of a DNA sequence that will eventually be converted into protein (i.e. the coding part).
- Long interspersed elements (LINEs): a subclass of retrotransposons that encode their own DNA- cutting exonucleases, RTases, and nuclear chaperone proteins.
- LINE-1 (L1): the most abundant family of LINEs. Most are not functional.
- Messenger RNA (mRNA): an intermediate in the central dogma of biology. Enable DNA messages to be converted into proteins or functional molecules the cell can use as enzymes, antibodies, hormones, or much more.
- RNA Polymerase II: an enzyme that makes a strand of mRNA from a DNA template.
- Human immunodeficiency virus (HIV): a retrovirus that targets immune cells, compromising our immune system and proliferating unnoticed and unchecked.
- APOBEC: a class of proteins that can inhibit the activity of RTases.
- Viral infectivity factor (Vif): a protein that prevents APOBEC proteins from inhibiting RTases.
- Intronic DNA: DNA that is spliced out of what will become a mature mRNA sequence ready for translation. It is not converted into protein.
- Epigenetics: The study of changes in organisms caused by modification of gene expression as opposed to modification of the actual DNA sequence.
Comments
Post a Comment